07.04.2015 13.10.2015
DNK (deoksiribonukleinska kiselina) je makromolekula koja prenosi informacije o tijelu s jedne generacije na drugu.
Proteini formiraju polipeptidne lance, informacije o kojima pohranjuje deoksiribonukleinska kiselina. Svaki region koji sadrži podatke o takvom lancu naziva se gen. Molekuli dezoksiribonukleinske kiseline, smješteni unutar jedne ćelije, zajedno predstavljaju nosioca genetske informacije o cijelom organizmu.
Otkriće molekule deoksiribonukleinske kiseline napravljeno je davne 1869. godine. Švicarski fiziolog Friedrich Miescher otkrio je supstancu koju je nazvao nuklein. Značaj velikog otkrića u početku nije bio cijenjen onako kako je trebao biti. Dugo se vjerovalo da nuklein nije ništa drugo do skladište fosfora.
Dolaskom 20. vijeka nastavljeno je proučavanje deoksiribonukleinske kiseline, međutim, početkom stoljeća velika većina naučnika u ovoj oblasti nije ni zamišljala da je DNK prenosilac informacija. Po njihovom mišljenju, njegova struktura je previše jednostavna i repetitivna da bi imala tako složenu funkciju.
Naučni proboj se dogodio 1944. godine kada je utvrđeno da je DNK od velikog naučnog značaja. Naučnik Oswald Avery, zajedno sa dvojicom kolega McLean McCarthy i Colin MacLeod, proveo je istraživanje o deoksiribonukleinskoj kiselini, što je rezultiralo publikacijom u The Journal of Experimental Medicine. U članku je dokazano da je deoksiribonukleinska kiselina „materijal“ gena i nosilac nasljednih informacija.
Čim je dokazano da deoksiribonukleinska kiselina nije ništa drugo do genski kod organizma i da ima važnu ulogu kao nosilac informacija, istraživanja biologa krenula su u pravom smjeru. Počelo je brzo proučavanje kola i interkonekcija. Do 1950. godine bilo je moguće samo utvrditi da se molekula DNK sastoji od nuklidnih lanaca, ali kako su oni međusobno povezani i koliko ih je ostalo nepoznato.
Tek 1953. godine utvrđeno je da unutar molekula deoksiribonukleinske kiseline postoje odnosi između različitih tipova azotnih baza. Sama molekula DNK bila je predstavljena kao dvostruka spirala.
Prijenos nasljednih informacija dezoksiribonukleinskom kiselinom može se uporediti s načinom na koji ljudi razmjenjuju informacije. To radimo pomoću zvukova i slova. U DNK se koriste baze dušične kiseline.
Svaka spirala makromolekula sastoji se od azotnih baza, ribbonske kiseline i ostatka fosforne kiseline. Karike mogu imati različite sekvence, njihova glavna karakteristika je da su sve usko povezane sa nizom druge spirale. Ovo svojstvo se naziva pravilo komplementarnosti.
Dvostruka spirala polimernih lanaca je poput merdevina od užeta. Svaki korak u njemu je par nukleotida koji je povezan jedinjenjem šećera i fosfata. Glavna razlika između molekula DNK i drugih je redoslijed parova. Ali upravo je taj raspored šifra prema kojoj se određuje redoslijed proteina koje stanice proizvode.
 Upoređujući proces sa ljudskim tipom nosioca i prenosa informacija, možemo reći da je u ovom slučaju riječ o lošoj azbuci, u kojoj postoje samo četiri slova. Sve riječi, kao i rečenice, sastavljene su od njih.
Upoređujući proces sa ljudskim tipom nosioca i prenosa informacija, možemo reći da je u ovom slučaju riječ o lošoj azbuci, u kojoj postoje samo četiri slova. Sve riječi, kao i rečenice, sastavljene su od njih.
Šifra je dešifrovana kada su ljudi shvatili da kod nije binarni, već trostruki. Svaka aminokiselina u proteinu apsolutno odgovara sekvenci od tri nukleotida u RNK i DNK, kodonima.
Deoksiribonukleinska kiselina prenosi informacije dva puta: kada se dijeli na dva dijela i kada kodira protein. Tako se podaci prenose u novoformiranu ćeliju. Tokom procesa replikacije, DNK pravi kopiju sebe. Pramenovi koji povezuju spiralu se razdvajaju i stvara se novi komplementarni lanac. Svaka od dvije novoformirane ćelije sadrži identičnu kopiju deoksiribonukleinske kiseline. Tako su sačuvane sve genetske informacije.
Stečeno znanje o molekulu DNK teško je precijeniti. Njihova praktična primjena je od velike važnosti za čovječanstvo. U suštini, otkrivanjem tajne makromolekula, ljudi su dobili pristup genima. Razvoj nauke o deoksiribonukleinskoj kiselini otvara neograničene mogućnosti za biologiju i medicinu.
Saznanja o nasljednoj prirodi deoksiribonukleinske kiseline našla su praktičnu primjenu u genetskom inženjeringu, što utiče na razvoj kliničke medicine. Metode zasnovane na proučavanju rekombinantne DNK otvorile su nove mogućnosti za proučavanje nasljednih bolesti.
Tehnologija koja se koristi za rekombinantne DNK molekule bila je revolucionarna za nauku proučavanja živih ćelija. Medicini i industriji otvorili su se novi načini da dobiju dovoljne količine onih proteina koji su se ranije dobijali u ograničenim količinama ili ih uopšte nisu dobijali.
Nažalost, istraživanje je daleko od završenog. Međutim, do danas je urađeno mnogo. To su metode kloniranja DNK i genetski inženjering. Tehnologija rekombinantne DNK postala je pravi proboj u medicini. Omogućava transplantaciju genetskog materijala iz jednog organizma u drugi. Smjer je u procesu proučavanja i razvoja, međutim, neki njegovi nalazi se već aktivno koriste u praksi.
Genska terapija je omogućila uvođenje potpuno zdravih gena u tijela bolesnih ljudi, sposobnih u potpunosti  rad. Ovo omogućava obnavljanje metaboličkih poremećaja uzrokovanih mutantnim genima. Danas se ova metoda koristi za liječenje djece sa imunodeficijencijom koja je uzrokovana defektom adenozin deaminaze.
rad. Ovo omogućava obnavljanje metaboličkih poremećaja uzrokovanih mutantnim genima. Danas se ova metoda koristi za liječenje djece sa imunodeficijencijom koja je uzrokovana defektom adenozin deaminaze.
Razvoj tretmana za mnoge bolesti upotrebom tehnologije rekombinantne DNK je u fazi kliničkog istraživanja. To su bolesti kao što su:
Hemofilija B, definisana prisustvom krvarenja tipa hematoma;
Porodična hiperholesterolemija;
Cistična fibroza itd.
Ako je genetika u procesu aktivnog razvoja u medicini, onda je ona dala najznačajnije praktične rezultate u poljoprivredi. Zahvaljujući njoj poljoprivredna proizvodnja je dostigla novi nivo. Razvijaju se nove sorte biljaka koje su od interesa za čovječanstvo. Zadatak naučnika nije samo da razviju nove sorte, već i da im usade najkorisnije kvalitete.
Nauka o deoksiribonukleinskoj kiselini se aktivno razvija, ali je unatoč tome još uvijek u početnoj fazi svog razvoja. Šta naučnici očekuju u završnoj fazi? Ovo je potpuna pobjeda nad fenomenima kao što su bolest i glad, te sposobnost kloniranja živih organizama i promjena karakteristika organizama. Možda će se uskoro razviti novi tip osobe, koji će biti savršena slika kojoj svi težimo kroz našu istoriju.
Razotkrivanje misterije DNK označilo je početak nove ere u razvoju biologije. Kako se proučavalo, dogodila su se ne samo naučna otkrića, već i zanimljivosti i zabavni slučajevi.
Na primjer, proučavajući voćne mušice, naučnici su svojim otkrićima počeli davati smiješna imena. Par gena koji dovode do odsustva spoljašnjih genitalnih organa kod ženki i muškaraca dobio je imena za lutke "Barbi" i "Ken", a mutantni gen, čiji vlasnik brzo umire, dobio je ime po čuvenom crtanom liku iz crtani film "Sousepark" "Kenny."
Naučnici proučavaju deoksiribonukleinsku kiselinu i rezultate istraživanja primjenjuju u praksi. Rezultati njihovog rada važni su za čovečanstvo. Genetičari i nauka koju promovišu imaju moć promijeniti svijet i učiniti ga boljim.
Reprodukcija svoje vrste jedno je od osnovnih svojstava živih bića. Zahvaljujući ovom fenomenu, postoji sličnost ne samo između organizama, već i između pojedinačnih ćelija, kao i njihovih organela (mitohondrija i plastida). Materijalna osnova ove sličnosti je prijenos genetskih informacija šifriranih u DNK nukleotidnom nizu, koji se odvija kroz procese DNK replikacije (samo-duplikacije). Sve karakteristike i svojstva ćelija i organizama ostvaruju se zahvaljujući proteinima, čija je struktura prvenstveno određena slijedom nukleotida DNK. Stoga, biosinteza nukleinskih kiselina i proteina igra od najveće važnosti u metaboličkim procesima. Strukturna jedinica nasljedne informacije je gen.
Nasljedna informacija u ćeliji nije monolitna, podijeljena je na zasebne “riječi” - gene.
Gene je elementarna jedinica genetske informacije.
Rad na programu “Ljudski genom”, koji je istovremeno sproveden u nekoliko zemalja i završen početkom ovog veka, dao nam je razumevanje da osoba ima samo oko 25-30 hiljada gena, ali informacije iz većine našeg DNK se nikada ne čita, jer sadrži ogroman broj besmislenih sekcija, ponavljanja i gena koji kodiraju osobine koje su izgubile značenje za ljude (rep, dlake na tijelu, itd.). Osim toga, dešifrovan je niz gena odgovornih za razvoj nasljednih bolesti, kao i ciljnih gena lijeka. Međutim, praktična primjena rezultata dobijenih tokom implementacije ovog programa odgađa se dok se ne dešifruju genomi većeg broja ljudi i ne postane jasno po čemu se razlikuju.
Geni koji kodiraju primarnu strukturu proteina, ribosomske ili transferne RNK se nazivaju strukturalni i geni koji omogućavaju aktivaciju ili supresiju čitanja informacija iz strukturnih gena - regulatorni. Međutim, čak i strukturni geni sadrže regulatorne regije.
Nasljedne informacije organizama su šifrirane u DNK u obliku određenih kombinacija nukleotida i njihovog niza - genetski kod. Njegova svojstva su: trostrukost, specifičnost, univerzalnost, redundantnost i nepreklapanje. Osim toga, u genetskom kodu nema znakova interpunkcije.
Svaka aminokiselina je kodirana u DNK sa tri nukleotida - trojka, na primjer, metionin je kodiran TAC tripletom, odnosno kod je triplet. S druge strane, svaki triplet kodira samo jednu aminokiselinu, što je njena specifičnost ili nedvosmislenost. Genetski kod je univerzalan za sve žive organizme, odnosno nasljedne informacije o ljudskim proteinima mogu čitati bakterije i obrnuto. Ovo ukazuje na jedinstvo porijekla organskog svijeta. Međutim, 64 kombinacije od tri nukleotida odgovaraju samo 20 aminokiselina, zbog čega jedna aminokiselina može biti kodirana sa 2-6 tripleta, odnosno genetski kod je suvišan ili degenerisan. Tri trojke nemaju odgovarajuće aminokiseline, nazivaju se stop kodoni, jer ukazuju na kraj sinteze polipeptidnog lanca.
Slijed baza u DNK tripletima i aminokiseline koje kodiraju
*Stop kodon, koji označava kraj sinteze polipeptidnog lanca.
Skraćenice za nazive aminokiselina:
Ala - alanin
Arg - arginin
Asn - asparagin
Asp - asparaginska kiselina
Val - valin
Njegov - histidin
Gly - glicin
Gln - glutamin
Glu - glutaminska kiselina
Ile - izoleucin
Leu - leucin
Liz - lizin
Met - metionin
Pro-prolin
Ser - serin
Tyr - tirozin
Tre - treonin
Tri-triptofan
Fen - fenilalanin
Cis - cistein
Ako počnete čitati genetske informacije ne od prvog nukleotida u tripletu, već od drugog, ne samo da će se okvir čitanja pomaknuti - protein sintetiziran na ovaj način bit će potpuno drugačiji ne samo u nukleotidnoj sekvenci, već i po strukturi i svojstva. Između trojki nema znakova interpunkcije, tako da nema prepreka za pomicanje okvira čitanja, što otvara prostor za nastanak i održavanje mutacija.
Bakterijske ćelije su sposobne da se udvostruče svakih 20-30 minuta, a eukariotske ćelije - svaki dan i još češće, što zahteva veliku brzinu i tačnost replikacije DNK. Osim toga, svaka stanica sadrži stotine i tisuće kopija mnogih proteina, posebno enzima, stoga je "parealni" način njihove proizvodnje neprihvatljiv za njihovu reprodukciju. Progresivnija metoda je žigosanje, koje vam omogućava da dobijete brojne točne kopije proizvoda i smanjite njegovu cijenu. Za štancanje je potrebna matrica iz koje se pravi otisak.
U ćelijama princip sinteze šablona je da se novi molekuli proteina i nukleinskih kiselina sintetišu u skladu sa programom ugrađenim u strukturu već postojećih molekula istih nukleinskih kiselina (DNK ili RNK).
DNK replikacija. DNK je dvolančani biopolimer, čiji su monomeri nukleotidi. Kada bi se biosinteza DNK odvijala na principu fotokopiranja, neminovno bi se pojavila brojna izobličenja i greške u nasljednim informacijama, što bi u konačnici dovelo do smrti novih organizama. Stoga se proces udvostručavanja DNK odvija drugačije, na polukonzervativan način: molekula DNK se odmotava i na svakom od lanaca se sintetiše novi lanac prema principu komplementarnosti. Proces samoreprodukcije molekula DNK, koji osigurava precizno kopiranje nasljednih informacija i njihov prijenos s generacije na generaciju, naziva se replikacija(od lat. replicationo- ponavljanje). Kao rezultat replikacije, formiraju se dvije apsolutno točne kopije molekula DNK majke, od kojih svaka nosi po jednu kopiju DNK majke.

Proces replikacije je zapravo izuzetno složen, budući da je u njemu uključen veliki broj proteina. Neki od njih odmotavaju dvostruku spiralu DNK, drugi razbijaju vodikove veze između nukleotida komplementarnih lanaca, treći (na primjer, enzim DNK polimeraza) odabiru nove nukleotide na principu komplementarnosti, itd. Dvije molekule DNK nastale kao rezultat replikacije se razilaze na dva tokom diobe novoformirane kćerke ćelije.
Greške u procesu replikacije nastaju izuzetno rijetko, ali ako se i dogode, vrlo brzo se eliminiraju kako DNK polimerazama tako i posebnim enzimima za popravak, jer svaka greška u nukleotidnoj sekvenci može dovesti do nepovratne promjene strukture i funkcija proteina. i, na kraju, negativno utiču na održivost nove ćelije ili čak pojedinca.
Biosinteza proteina. Kao što je figurativno rekao istaknuti filozof 19. veka F. Engels: „Život je oblik postojanja proteinskih tela“. Struktura i svojstva proteinskih molekula određuju se njihovom primarnom strukturom, odnosno sekvencom aminokiselina kodiranih u DNK. Od tačnosti reprodukcije ovih informacija zavisi ne samo postojanje samog polipeptida, već i funkcionisanje ćelije u celini, pa je proces sinteze proteina od velike važnosti. Čini se da je to najsloženiji proces sinteze u ćeliji, jer uključuje do tri stotine različitih enzima i drugih makromolekula. Osim toga, teče velikom brzinom, što zahtijeva još veću preciznost.
Postoje dvije glavne faze u biosintezi proteina: transkripcija i translacija.
Transkripcija(od lat. transkripcija- prepisivanje) je biosinteza mRNA molekula na DNK matrici.

Budući da molekula DNK sadrži dva antiparalelna lanca, čitanje informacija iz oba lanca dovelo bi do stvaranja potpuno različitih mRNA, pa je njihova biosinteza moguća samo na jednom od lanaca, koji se naziva kodirajući, odnosno kodogen, za razliku od drugog, nekodirajući ili nekodogeni. Proces ponovnog pisanja osigurava poseban enzim, RNA polimeraza, koji bira RNA nukleotide prema principu komplementarnosti. Ovaj proces se može dogoditi i u jezgri i u organelama koje imaju svoju DNK - mitohondrije i plastide.
Molekuli mRNA sintetizirani tokom transkripcije prolaze kroz složen proces pripreme za translaciju (mitohondrijalne i plastidne mRNA mogu ostati unutar organela, gdje se odvija druga faza biosinteze proteina). Tokom procesa sazrijevanja mRNA, za nju su vezana prva tri nukleotida (AUG) i rep adenil nukleotida, čija dužina određuje koliko kopija proteina može biti sintetizirano na datom molekulu. Tek tada zrele mRNA napuštaju jezgro kroz nuklearne pore.
Paralelno, u citoplazmi se odvija proces aktivacije aminokiselina, tokom kojeg se aminokiselina pridružuje odgovarajućoj slobodnoj tRNK. Ovaj proces katalizira poseban enzim i zahtijeva ATP.
Broadcast(od lat. emitovanje- transfer) je biosinteza polipeptidnog lanca na matrici mRNA, tokom koje se genetska informacija prevodi u aminokiselinsku sekvencu polipeptidnog lanca.
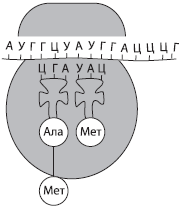
Druga faza sinteze proteina najčešće se javlja u citoplazmi, na primjer na grubom ER. Za njen nastanak neophodno je prisustvo ribozoma, aktivacija tRNK, pri čemu se vezuju odgovarajuće aminokiseline, prisustvo Mg2+ jona, kao i optimalni uslovi okoline (temperatura, pH, pritisak, itd.).
Za početak emitovanja ( iniciranje) mala ribosomalna podjedinica je vezana za molekul mRNA spreman za sintezu, a zatim se, prema principu komplementarnosti prvom kodonu (AUG), odabire tRNA koja nosi aminokiselinu metionin. Tek nakon toga se veže velika ribosomska podjedinica. Unutar sastavljenog ribozoma postoje dva kodona mRNA, od kojih je prvi već zauzet. Druga tRNA, koja također nosi aminokiselinu, dodaje se kodonu uz nju, nakon čega se uz pomoć enzima formira peptidna veza između aminokiselinskih ostataka. Ribosom pomiče jedan kodon mRNA; prva tRNA oslobođena aminokiseline vraća se u citoplazmu nakon sljedeće aminokiseline, a fragment budućeg polipeptidnog lanca visi, takoreći, na preostaloj tRNK. Sljedeća tRNA se veže za novi kodon koji se nađe unutar ribozoma, proces se ponavlja i korak po korak se polipeptidni lanac produžuje, tj. izduženje.
Kraj sinteze proteina ( raskid) se javlja čim se u molekulu mRNA naiđe na specifičnu sekvencu nukleotida koja ne kodira aminokiselinu (stop kodon). Nakon toga se odvajaju ribosom, mRNA i polipeptidni lanac, a novosintetizovani protein dobija odgovarajuću strukturu i transportuje se u deo ćelije gde će obavljati svoje funkcije.
Translacija je energetski vrlo intenzivan proces, budući da se energija jedne molekule ATP-a troši za pričvršćivanje jedne aminokiseline na tRNA, a još nekoliko se koristi za pomicanje ribozoma duž mRNA molekula.
Da bi se ubrzala sinteza određenih proteinskih molekula, nekoliko ribozoma može se sukcesivno vezati za molekulu mRNA, koji formiraju jednu strukturu - polizom.
Ćelija je genetska jedinica živog bića. Kromosomi, njihova struktura (oblik i veličina) i funkcije. Broj hromozoma i konstantnost njihove vrste. Somatske i zametne ćelije. Životni ciklus ćelije: interfaza i mitoza. Mitoza je podjela somatskih ćelija. Mejoza. Faze mitoze i mejoze. Razvoj zametnih stanica u biljkama i životinjama. Podjela ćelija je osnova za rast, razvoj i reprodukciju organizama. Uloga mejoze i mitoze
Unatoč činjenici da su nukleinske kiseline nosilac genetske informacije, implementacija tih informacija je nemoguća izvan ćelije, što se lako dokazuje na primjeru virusa. Ovi organizmi, koji često sadrže samo DNK ili RNK, ne mogu se razmnožavati nezavisno da bi to učinili, moraju koristiti nasljedni aparat ćelije. Ne mogu čak ni prodrijeti u ćeliju bez pomoći same ćelije, osim upotrebom membranskih transportnih mehanizama ili zbog oštećenja ćelije. Većina virusa je nestabilna; oni umiru nakon samo nekoliko sati izlaganja na otvorenom. Shodno tome, ćelija je genetska jedinica živog bića, koja ima minimalan skup komponenti za očuvanje, promjenu i implementaciju nasljednih informacija, kao i njihovo prenošenje na potomke.
Većina genetskih informacija eukariotske ćelije nalazi se u jezgru. Posebnost njegove organizacije je da, za razliku od DNK prokariotske ćelije, molekuli DNK eukariota nisu zatvoreni i formiraju složene komplekse s proteinima - hromozomima.
hromozom(iz grčkog hrom- boja, boja i soma- tijelo) je struktura ćelijskog jezgra, koja sadrži gene i nosi određene nasljedne informacije o karakteristikama i svojstvima organizma.
Ponekad se kružni DNK molekuli prokariota nazivaju i hromozomi. Kromosomi su sposobni za samoumnožavanje, imaju strukturnu i funkcionalnu individualnost i zadržavaju je generacijama. Svaka ćelija nosi sve nasljedne informacije tijela, ali samo mali dio radi u njoj.
Osnova hromozoma je dvolančana DNK molekula prepuna proteina. Kod eukariota, histonski i nehistonski proteini stupaju u interakciju sa DNK, dok kod prokariota histonski proteini odsutni.
Hromozomi se najbolje vide pod svjetlosnim mikroskopom tokom diobe ćelije, kada kao rezultat zbijanja poprimaju izgled štapićastih tijela razdvojenih primarnim suženjem - centromere - na ramenima. Može biti i na hromozomu sekundarna konstrikcija, koji u nekim slučajevima razdvaja tzv satelit. Zovu se krajevi hromozoma telomere. Telomeri sprečavaju da se krajevi hromozoma lepe zajedno i obezbeđuju njihovo pričvršćivanje za nuklearnu membranu u ćeliji koja se ne deli. Na početku diobe hromozomi se udvostručuju i sastoje se od dva ćerka hromozoma - hromatida, pričvršćen na centromeri.

Prema svom obliku, hromozomi se dijele na ravnokrake, nejednakokrake i štapićaste hromozome. Veličine hromozoma značajno variraju, ali prosječni hromozom ima dimenzije od 5 $×$ 1,4 mikrona.
U nekim slučajevima, hromozomi, kao rezultat brojnih duplikacija DNK, sadrže stotine i hiljade hromatida: takvi se ogromni hromozomi nazivaju polytene. Nalaze se u pljuvačnim žlijezdama larvi Drosophila, kao i u probavnim žlijezdama okruglih crva.
Prema ćelijskoj teoriji, ćelija je jedinica strukture, vitalne aktivnosti i razvoja organizma. Dakle, tako važne funkcije živih bića kao što su rast, reprodukcija i razvoj organizma se obezbjeđuju na ćelijskom nivou. Ćelije višećelijskih organizama mogu se podijeliti na somatske i reproduktivne stanice.
Somatske ćelije- sve su to ćelije tijela nastale kao rezultat mitotičke diobe.
Proučavanje hromozoma omogućilo je da se utvrdi da se somatske ćelije tijela svake biološke vrste karakteriziraju konstantnim brojem kromosoma. Na primjer, osoba ih ima 46. Skup hromozoma somatskih ćelija se zove diploidni(2n), ili duplo.

Polne ćelije, ili gamete, su specijalizovane ćelije koje se koriste za seksualnu reprodukciju.
Gamete uvijek sadrže upola manje hromozoma od somatskih ćelija (kod ljudi - 23), pa se skup hromozoma zametnih ćelija naziva haploidni(n) ili pojedinac. Njegovo stvaranje povezano je s mejotskom diobom stanica.
Količina DNK u somatskim ćelijama označena je kao 2c, au polnim ćelijama - 1c. Genetska formula somatskih ćelija je zapisana kao 2n2c, a polnih ćelija - 1n1c.
U jezgrima nekih somatskih stanica broj kromosoma može se razlikovati od broja u somatskim stanicama. Ako je ta razlika veća od jednog, dva, tri, itd. haploidnih skupova, tada se takve ćelije nazivaju poliploid(tri-, tetra-, pentaploid, respektivno). U takvim ćelijama se metabolički procesi obično odvijaju vrlo intenzivno.
Broj hromozoma sam po sebi nije specifično obilježje vrste, budući da različiti organizmi mogu imati jednak broj hromozoma, ali srodni mogu imati različit broj. Na primjer, malarijski plazmodijum i konjski okrugli crv imaju po dva hromozoma, dok ljudi i čimpanze imaju 46, odnosno 48.
Ljudski hromozomi se dele u dve grupe: autozomi i polni hromozomi (heterohromozomi). Autosome u ljudskim somatskim ćelijama ima 22 para, isti su za muškarce i žene, i polni hromozomi samo jedan par, ali to je ono što određuje pol pojedinca. Postoje dvije vrste polnih hromozoma - X i Y. Ženske tjelesne ćelije nose dva X hromozoma, a muške - X i Y.
Kariotip- ovo je skup karakteristika hromozomskog seta organizma (broj kromosoma, njihov oblik i veličina).
Uslovni zapis kariotipa uključuje ukupan broj hromozoma, polnih hromozoma i moguća odstupanja u setu hromozoma. Na primjer, kariotip normalnog muškarca piše se kao 46,XY, a kariotip normalne žene kao 46,XX.
Ćelije ne nastaju svaki put iznova, one nastaju samo kao rezultat diobe matičnih stanica. Nakon diobe ćelijama kćeri potrebno je neko vrijeme da formiraju organele i steknu odgovarajuću strukturu koja bi osigurala obavljanje određene funkcije. Ovaj vremenski period se zove sazrevanje.
Naziva se vremenski period od pojave ćelije kao rezultat diobe do njene diobe ili smrti životni ciklus ćelije.
U eukariotskim ćelijama životni ciklus je podijeljen u dvije glavne faze: interfazu i mitozu.
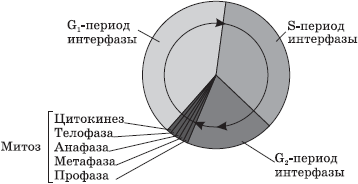
Interfaza- ovo je vremenski period u životnom ciklusu tokom kojeg se ćelija ne dijeli i normalno funkcionira. Interfaza je podijeljena na tri perioda: G 1 -, S- i G 2 -periodi.
G 1 -period(presintetički, postmitotski) je period rasta i razvoja ćelije tokom kojeg se odvija aktivna sinteza RNK, proteina i drugih supstanci neophodnih za potpuni život novonastale ćelije. Pred kraj ovog perioda, ćelija može početi da se priprema za dupliciranje svoje DNK.
IN S-period(sintetički) dolazi do samog procesa replikacije DNK. Jedini dio hromozoma koji ne podliježe replikaciji je centromera, pa se tako nastale DNK molekule ne divergiraju u potpunosti, već se drže zajedno u njemu, a na početku diobe hromozom ima X-oblik. Genetska formula ćelije nakon udvostručenja DNK je 2n4c. Takođe u S-periodu, centrioli ćelijskog centra su udvostručeni.
G 2 -period(postsintetski, premitotički) karakteriše intenzivna sinteza RNK, proteina i ATP-a neophodnih za proces deobe ćelije, kao i odvajanje centriola, mitohondrija i plastida. Do kraja interfaze, kromatin i nukleolus ostaju jasno razlučivi, integritet nuklearnog omotača nije narušen, a organele se ne mijenjaju.
Neke od tjelesnih stanica su u stanju da obavljaju svoje funkcije tokom cijelog života u tijelu (neuroni našeg mozga, mišićne ćelije srca), dok druge postoje kratko, nakon čega odumiru (ćelije crijevnog epitela, epidermalne ćelije koža). Shodno tome, tijelo mora stalno prolaziti kroz procese diobe stanica i formiranja novih koje bi zamijenile mrtve. Zovu se ćelije sposobne za dijeljenje stablo. U ljudskom tijelu nalaze se u crvenoj koštanoj srži, u dubokim slojevima epiderme kože i na drugim mjestima. Koristeći ove ćelije, možete uzgajati novi organ, postići podmlađivanje, a također i klonirati tijelo. Izgledi za korištenje matičnih stanica su apsolutno jasni, ali se još uvijek raspravlja o moralnim i etičkim aspektima ovog problema, jer se u većini slučajeva koriste embrionalne matične stanice dobivene iz ljudskih embriona ubijenih tijekom pobačaja.
Trajanje interfaze u biljnim i životinjskim ćelijama je u prosjeku 10-20 sati, dok mitoza traje oko 1-2 sata.
Tokom uzastopnih podjela u višećelijskim organizmima, ćelije kćeri postaju sve raznovrsnije jer čitaju informacije iz sve većeg broja gena.
Neke stanice se s vremenom prestaju dijeliti i umiru, što može biti zbog dovršetka određenih funkcija, kao u slučaju epidermalnih stanica kože i krvnih stanica, ili zbog oštećenja ovih stanica faktorima okoline, posebno patogenima. Genetski programirana ćelijska smrt se naziva apoptoza, dok nesrećna smrt - nekroza.
Mitoza- metoda indirektne diobe somatskih stanica.
Tokom mitoze, ćelija prolazi kroz niz uzastopnih faza, usled čega svaka ćerka ćelija dobija isti set hromozoma kao i matična ćelija.
Mitoza je podijeljena u četiri glavne faze: profaza, metafaza, anafaza i telofaza. Profaza- najduža faza mitoze, tokom koje se hromatin kondenzuje, što rezultira hromozomima u obliku slova X koji se sastoje od dve hromatide (kromosoma kćeri) postaju vidljivi. U tom slučaju nukleolus nestaje, centriole se razilaze prema polovima stanice i počinje se formirati akromatinsko vreteno (diobno vreteno) iz mikrotubula. Na kraju profaze, nuklearna membrana se raspada u zasebne vezikule.

IN metafaza Kromosomi su poređani duž ekvatora ćelije sa svojim centromerama, za koje su pričvršćene mikrotubule potpuno formiranog vretena. U ovoj fazi podjele, kromosomi su najkompaktniji i imaju karakterističan oblik, što omogućava proučavanje kariotipa.
IN anafaza Brza replikacija DNK događa se na centromerama, zbog čega se hromozomi cijepaju i hromatide divergiraju do polova ćelije, rastegnute mikrotubulama. Raspodjela hromatida mora biti apsolutno jednaka, jer upravo ovaj proces osigurava održavanje konstantnog broja kromosoma u stanicama tijela.
Na sceni telofaze kćeri hromozomi se skupljaju na polovima, oko njih se iz vezikula formiraju despiralne, nuklearne membrane, a u novonastalim jezgrama pojavljuju se jezgre.
Nakon nuklearne diobe dolazi do diobe citoplazme - citokineza, tokom koje dolazi do manje ili više ujednačene raspodele svih organela matične ćelije.
Dakle, kao rezultat mitoze, iz jedne matične ćelije nastaju dvije kćerke ćelije, od kojih je svaka genetska kopija matične ćelije (2n2c).
U bolesnim, oštećenim, ostarjelim stanicama i specijalizovanim tkivima tijela može se dogoditi nešto drugačiji proces diobe - amitoza. Amitoza naziva se direktna dioba eukariotskih stanica, u kojoj ne dolazi do stvaranja genetski ekvivalentnih stanica, jer su ćelijske komponente neravnomjerno raspoređene. Nalazi se u biljkama u endospermu, a kod životinja - u jetri, hrskavici i rožnici oka.
Mejoza je metoda indirektne diobe primarnih zametnih stanica (2n2c), koja rezultira stvaranjem haploidnih stanica (1n1c), najčešće zametnih stanica.
Za razliku od mitoze, mejoza se sastoji od dvije uzastopne diobe stanica, od kojih svakoj prethodi interfaza. Prva podjela mejoze (mejoza I) se zove redukcionistički, budući da je u ovom slučaju broj hromozoma prepolovljen, a druga podjela (mejoza II) - equational, budući da je u njegovom procesu sačuvan broj hromozoma.

Interfaza I nastavlja kao interfaza mitoze. Mejoza I podijeljen je u četiri faze: profaza I, metafaza I, anafaza I i telofaza I. B profaza I Događaju se dva važna procesa - konjugacija i krosing. Konjugacija- Ovo je proces fuzije homolognih (uparenih) hromozoma cijelom dužinom. Parovi hromozoma nastali tokom konjugacije čuvaju se do kraja metafaze I.
Prelazak- međusobna izmjena homolognih regija homolognih hromozoma. Kao rezultat križanja, hromozomi koje tijelo primi od oba roditelja dobivaju nove kombinacije gena, što uzrokuje pojavu genetski raznolikog potomstva. Na kraju profaze I, kao iu profazi mitoze, jezgro nestaje, centriole se divergiraju prema polovima ćelije, a nuklearna membrana se raspada.

IN metafaza I parovi hromozoma poredani su duž ekvatora ćelije, a mikrotubule vretena su pričvršćene za njihove centromere.
IN anafaza I Cijeli homologni hromozomi, koji se sastoje od dvije hromatide, divergiraju do polova.
IN telofaza I Nuklearne membrane se formiraju oko klastera hromozoma na polovima ćelije i formiraju se jezgre.
Citokineza I osigurava odvajanje citoplazme ćelija kćeri.
Ćerke ćelije (1n2c) nastale kao rezultat mejoze I su genetski heterogene, budući da njihovi hromozomi, nasumično raspoređeni na polove ćelije, sadrže različite gene.
Komparativne karakteristike mitoze i mejoze
| Potpiši | Mitoza | Mejoza | |
| Koje ćelije počinju da se dele? | somatski (2n) | Primarne zametne ćelije (2n) | |
| Broj divizija | 1 | 2 | |
| Koliko i kakvih ćelija se formira tokom deobe? | 2 somatske (2n) | 4 seksualne (n) | |
| Interfaza | Priprema ćelije za podelu, udvostručavanje DNK | Vrlo kratko, ne dolazi do udvostručavanja DNK | |
| Faze | Mejoza I | Mejoza II | |
| Profaza | Može doći do kondenzacije hromozoma, nestanka nukleola, dezintegracije nuklearne membrane, konjugacije i krosingovera | Kondenzacija hromozoma, nestanak nukleola, dezintegracija nuklearne membrane | |
| Metafaza | Parovi hromozoma nalaze se duž ekvatora, formira se vreteno | Hromozomi se nižu duž ekvatora, formira se vreteno | |
| Anafaza | Homologni hromozomi iz dve hromatide kreću se prema polovima | Kromatide se kreću prema polovima | |
| Telofaza | Kromosomi su despiralni, formiraju se nove nuklearne membrane i jezgre | Kromosomi su despiralni, formiraju se nove nuklearne membrane i jezgre | |
Interfaza II vrlo kratko, pošto u njemu ne dolazi do udvostručenja DNK, odnosno nema S-perioda.
Mejoza II također podijeljen u četiri faze: profaza II, metafaza II, anafaza II i telofaza II. IN profaza II isti procesi se dešavaju kao u profazi I, sa izuzetkom konjugacije i krosingovera.
IN metafaza II hromozomi se nalaze duž ekvatora ćelije.
IN anafaza II hromozomi su podijeljeni na centromerima i hromatide su rastegnute prema polovima.
IN telofaza II Nuklearne membrane i jezgre formiraju se oko klastera kćeri hromozoma.
Poslije citokineza II Genetska formula sve četiri ćelije kćeri je 1n1c, ali sve imaju drugačiji skup gena, što je rezultat ukrštanja i slučajne kombinacije hromozoma majčinog i očinskog organizma u ćelijama kćeri.
Gametogeneza(iz grčkog gameta- supruga, gamete- muž i geneza- nastanak, nastanak) je proces formiranja zrelih zametnih ćelija.
Budući da su za seksualnu reprodukciju najčešće potrebne dvije jedinke - ženka i mužjak, koji proizvode različite polne stanice - jajašca i spermu, procesi formiranja ovih gameta moraju biti različiti.
Priroda procesa umnogome ovisi o tome da li se događa u biljnoj ili životinjskoj ćeliji, budući da se kod biljaka javlja samo mitoza prilikom formiranja gameta, a kod životinja i mitoza i mejoza.
Razvoj zametnih ćelija u biljkama. Kod kritosjemenjača formiranje muških i ženskih reproduktivnih stanica događa se u različitim dijelovima cvijeta - prašnicima i tučkom.
Prije formiranja muških reproduktivnih stanica - mikrogametogeneza(iz grčkog mikros- mali) - dešava se mikrosporogeneza, odnosno stvaranje mikrospora u prašnicima prašnika. Ovaj proces je povezan s mejotičkom diobom matične stanice, što rezultira četiri haploidne mikrospore. Mikrogametogeneza je povezana s mitotičkom podjelom mikrospore, pri čemu nastaje muški gametofit od dvije ćelije - velike vegetativno(sifonogena) i plitka generativno. Nakon diobe, muški gametofit postaje prekriven gustim membranama i formira polenovo zrno. U nekim slučajevima, čak i tokom procesa sazrevanja polena, a ponekad tek nakon prelaska na stigmu tučka, generativna ćelija se mitotički deli i formira dve nepokretne muške zametne ćelije - sperma. Nakon oprašivanja, iz vegetativne ćelije se formira polenova cijev kroz koju spermatozoidi prodiru u jajnik tučka radi oplodnje.

Razvoj ženskih zametnih ćelija u biljkama naziva se megagametogeneza(iz grčkog megas- veliki). Javlja se u jajniku tučka, kojem prethodi megasporogeneza, kao rezultat čega se mejotičkom diobom formiraju četiri megaspore iz matične stanice megaspore koja leži u nucelusu. Jedna od megaspora se mitotički dijeli tri puta, dajući ženskom gametofitu - embrionsku vrećicu sa osam jezgara. Naknadnim odvajanjem citoplazme ćelija kćeri, jedna od nastalih ćelija postaje jaje, na čijim stranama leže takozvani sinergidi, na suprotnom kraju embrionalne vrećice formiraju se tri antipoda, au centru , kao rezultat fuzije dva haploidna jezgra, nastaje diploidna centralna ćelija.

Razvoj zametnih ćelija kod životinja. Kod životinja postoje dva procesa formiranja zametnih ćelija - spermatogeneza i oogeneza.
Spermatogeneza(iz grčkog spermatozoida, spermatozoida- seme i geneza- porijeklo, pojava) je proces formiranja zrelih muških zametnih stanica - spermatozoida. Kod ljudi se javlja u testisima, odnosno testisima, i dijeli se na četiri perioda: razmnožavanje, rast, sazrijevanje i formiranje.

IN sezona razmnožavanja primordijalne zametne stanice dijele se mitotički, što rezultira stvaranjem diploida spermatogonije. IN period rasta spermatogonije akumuliraju hranjive tvari u citoplazmi, povećavaju se i pretvaraju u primarnih spermatocita, ili Spermatociti 1. reda. Tek nakon toga ulaze u mejozu ( period sazrevanja), zbog čega se prvo formiraju dva sekundarni spermatocit, ili Spermatocit 2. reda, a zatim - četiri haploidne ćelije sa još prilično velikom količinom citoplazme - spermatida. IN period formiranja gube gotovo svu svoju citoplazmu i formiraju flagellum, pretvarajući se u spermu.
Sperma, ili živahno, - vrlo male pokretne muške reproduktivne stanice s glavom, vratom i repom.

IN glava, pored jezgre, je akrozom- modificirani Golgijev kompleks, koji osigurava otapanje jajnih membrana tokom procesa oplodnje. IN cerviksa su centrioli ćelijskog centra i baze konjski rep formiraju mikrotubule koje direktno podržavaju kretanje spermatozoida. Sadrži i mitohondrije, koje spermiju obezbjeđuju ATP energiju za kretanje.
Oogeneza(iz grčkog UN- jaje i geneza- porijeklo, pojava) je proces formiranja zrelih ženskih zametnih stanica - jaja. Kod ljudi se javlja u jajnicima i sastoji se od tri perioda: reprodukcije, rasta i sazrijevanja. Periodi razmnožavanja i rasta, slični onima u spermatogenezi, javljaju se tokom intrauterinog razvoja. U ovom slučaju, diploidne ćelije nastaju iz primarnih zametnih ćelija kao rezultat mitoze. oogonia, koji se zatim pretvaraju u diploidne primarne oociti, ili Oociti 1. reda. Mejoza i naknadna citokineza koja se javlja u period sazrevanja, karakteriše neravnomerna podela citoplazme matične ćelije, tako da se kao rezultat toga dobija prvo sekundarne oocite, ili Oocita 2. reda, And prvo polarno tijelo, a zatim iz sekundarne oocite - jajeta, koje zadržava cjelokupnu zalihu hranjivih tvari, i drugo polarno tijelo, dok je prvo polarno tijelo podijeljeno na dva. Polarna tijela preuzimaju višak genetskog materijala.
Kod ljudi se jaja stvaraju u intervalima od 28-29 dana. Ciklus povezan sa sazrevanjem i oslobađanjem jajnih ćelija naziva se menstrualni.
Jaje- velika ženska reproduktivna stanica koja nosi ne samo haploidni skup hromozoma, već i značajnu zalihu hranjivih tvari za kasniji razvoj embrija.

Jaje kod sisara je prekriveno sa četiri membrane, koje smanjuju mogućnost oštećenja raznim faktorima. Promjer jajeta kod ljudi doseže 150-200 mikrona, dok u noja može biti nekoliko centimetara.
Ako kod jednoćelijskih organizama dioba stanica dovodi do povećanja broja jedinki, odnosno reprodukcije, onda kod višećelijskih organizama ovaj proces može imati različita značenja. Dakle, podjela embrionalnih stanica, počevši od zigote, predstavlja biološka osnova međusobno povezanih procesa rasta i razvoja. Slične promjene se primjećuju kod ljudi tokom adolescencije, kada se broj ćelija ne samo povećava, već se javlja i kvalitativna promjena u tijelu. Osnova reprodukcije višećelijskih organizama je i dioba ćelija, na primjer, kod aseksualne reprodukcije, zahvaljujući ovom procesu, obnavlja se cijeli dio organizma, a kod spolne reprodukcije, u procesu gametogeneze, nastaju polne ćelije, koji kasnije stvaraju novi organizam. Treba napomenuti da glavne metode diobe eukariotske ćelije - mitoza i mejoza - imaju različita značenja u životnim ciklusima organizama.
Kao rezultat mitoze, dolazi do ravnomjerne raspodjele nasljednog materijala između stanica kćeri - točne kopije majke. Bez mitoze, postojanje i rast višećelijskih organizama koji se razvijaju iz jedne ćelije, zigote, bili bi nemogući, jer sve ćelije takvih organizama moraju sadržavati iste genetske informacije.
U procesu diobe ćelije kćeri postaju sve raznovrsnije po strukturi i funkcijama, što je povezano sa aktivacijom sve više novih grupa gena u njima zbog međustanične interakcije. Dakle, mitoza je neophodna za razvoj organizma.
Ova metoda diobe ćelija neophodna je za procese aseksualne reprodukcije i regeneracije (obnove) oštećenih tkiva, kao i organa.
Mejoza, zauzvrat, osigurava postojanost kariotipa tijekom seksualne reprodukcije, budući da prepolovi skup kromosoma prije seksualne reprodukcije, koji se zatim obnavlja kao rezultat oplodnje. Osim toga, mejoza dovodi do pojave novih kombinacija roditeljskih gena zbog križanja i nasumične kombinacije hromozoma u ćelijama kćerima. Zahvaljujući tome, potomci su genetski raznoliki, što daje materijal za prirodnu selekciju i predstavlja materijalnu osnovu za evoluciju. Promjena broja, oblika i veličine hromozoma, s jedne strane, može dovesti do pojave raznih odstupanja u razvoju organizma, pa čak i njegove smrti, as druge strane može dovesti do pojave jedinki. prilagođeniji okruženju.
Dakle, ćelija je jedinica rasta, razvoja i reprodukcije organizama.
[email protected] u kategoriji, pitanje otvoreno 21.08.2017 u 18:41
A i RNK
B t RNA
U DNK
G hromozomi
Osnova individualnosti i specifičnosti organizama je:
I struktura tjelesnih proteina
B ćelijska struktura
U funkciji ćelije
D struktura aminokiselina
DNK nosi informacije o strukturi
I proteini, masti, ugljikohidrati
B proteini i masti
B aminokiseline
G proteini
Jedan gen kodira informacije:
I o strukturi nekoliko proteina
B o strukturi jednog od lanaca DNK
U primarnoj strukturi jednog proteinskog molekula
O strukturi aminokiselina
Koji nukleotid nije dio DNK?
I timin
Buracil
Za guanin
G citozin
D adenin
Koje se veze prekidaju u molekulu DNK kada se udvostruči?
I peptid
B kovalentna, između ugljikohidrata i fosfata
U vodoniku, između dva navoja
G ion
Koliko novih jednostrukih lanaca se sintetizira kada se jedan molekul udvostruči?
I četiri
B dva
U jednom
G tri
Koja je shema umnožavanja DNK tačna?
A kada se molekul DNK udvostruči, formira se potpuno novi ćerki molekul
B kćerka molekula DNK sastoji se od jednog starog i jednog novog lanca
Majčinska DNK se raspada na male fragmente, koji se zatim sklapaju u nove molekule kćeri
Koja činjenica potvrđuje da je DNK genetski materijal?
A količina DNK u ćelijama jednog organizma je konstantna
B DNK se sastoji od nukleotida
DNK je lokalizirana u ćelijskom jezgru
DNK je dvostruka spirala
Koja od sljedećih ljudskih ćelija ne sadrži DNK?
Zreli leukocit
B zrela crvena krvna zrnca
U limfocitima
G neuron
Ako je nukleotidni sastav DNK ATT-GCH-TAT, kakav bi onda trebao biti nukleotidni sastav mRNA?
A TAA-TsGTs-UTA
B TAA-GTG-UTU
U UAA-TsGTs-AUA
G UAA-TsGTs-ATA
Transkripcija se zove:
I proces formiranja Irka
B Proces umnožavanja DNK
Tokom formiranja proteinskog lanca na ribosomima
D proces spajanja tRNA sa aminokiselinama
Sinteza mRNA počinje:
I od razdvajanja molekula DNK na dva lanca
B sa udvostručavanjem svake niti
Interakcija između RNA polimeraze i gena
G sa cijepanjem gena na nukleotide
Aminokiselina triptofan je kodirana UGG. Koji DNK triplet nosi informacije o ovoj kiselini?
ACC
B TCC
U UCC
Gdje se mRNA sintetiše?
I u ribosomima
B u citoplazmi
U nukleolu
G u jezgru
Kako će izgledati dio lanca mRNA ako se drugi nukleotid prvog tripleta u DNK (GCT-AGT-CCA) zamijeni nukleotidom T?
A TsGA-UCA-GGT
B CAA-UCA-GGU
U GAU-ASU-CCA
G TsCU-UTSU-GGU
Ako šifra nije tri, već četiri slova, koliko bi se onda kombinacija u ovom slučaju moglo napraviti od četiri nukleotida?
A 4(4)
B 4(16)
B 2(4)
G 16(3)
Koje informacije sadrži jedan DNK triplet?
I informacije o redoslijedu aminokiselina u proteinu
B informacija o jednoj osobini organizma
Informacije o jednoj aminokiselini uključenoj u proteinski lanac
D informacija o početku sinteze mRNA
Koji enzim sintetiše mRNA?
RNA sintetaza
B RNA polimeraza
B DNK polimeraza
Dezoksiribonukleinska kiselina je nosilac nasledne informacije u ćeliji i sadrži deoksiribozu kao komponentu ugljenih hidrata, adenin (A), gvanin (G), citozin (C) i timin (T) kao azotne baze, kao i ostatak fosforne kiseline.
Rice. 12.
Sve ove strukture su formirane od dva antiparalelna DNK lanca koji se drže zajedno komplementarnim nukleotidnim parom. Svaki oblik je prikazan sa strane i odozgo. Šećerno-fosfatni parovi i bazni parovi su istaknuti u različitim nijansama sive: tamno sivoj i svijetlosivoj.
A. B-oblik DNK, koji se najčešće nalazi u ćelijama.
B. A-oblik DNK, koji postaje dominantan kada se bilo koja DNK osuši, bez obzira na njen slijed. B. Z-oblik DNK: neke sekvence dobijaju ovaj oblik pod određenim uslovima. B-forma i A-forma su desnoruki, a Z-oblik je ljevoruk (prema Albertsu).
DNK je dugačak, nerazgranati polimer koji se sastoji od samo četiri podjedinice - deoksiribonukleotida. Nukleotidi su međusobno povezani kovalentnim fosfodiestarskim vezama, povezujući 5" atom ugljika jednog ostatka sa 3" atomom ugljika sljedećeg ostatka. Četiri vrste baza su "nanizane" na lanac šećer-fosfata, kao četiri različite vrste perli nanizanih na jednu nit. Dakle, molekule DNK sastoje se od dva duga, komplementarna lanca koji se drže zajedno uparivanje baza.
Model DNK, prema kojem se sve baze DNK nalaze unutar dvostruke spirale, a šećerno-fosfatna kičma izvana, predložili su 1953. Watson i Crick. Broj efektivnih vodoničnih veza koje se mogu formirati između G i C ili između A i T će u ovom slučaju biti veći nego kod bilo koje druge kombinacije. Upravo je DNK model koji su predložili Watson i Crick omogućio da se formuliraju osnovni principi prijenosa nasljednih informacija zasnovanih na komplementarnosti dva lanca DNK. Jedan lanac služi kao predložak za formiranje njegovog komplementarnog lanca, a svaki nukleotid je slovo u abecedi od četiri slova.
Nukleotidi koji čine DNK sastoje se od cikličkog spoja koji sadrži dušik (azotna baza), ostatka šećera od pet ugljika i jedne ili više fosfatnih grupa. Glavna i najvažnija uloga nukleotida u ćeliji je da su oni monomeri od kojih se grade polinukleotidi - nukleinske kiseline odgovorne za skladištenje i prijenos bioloških informacija. Dvije glavne vrste nukleinskih kiselina razlikuju se po ostatku šećera u njihovoj polimernoj kičmi. Izgrađena na ribozi, ribonukleinska kiselina (RNA) sadrži adenin, gvanin, citozin i uracil. Deoksiribonukleinska kiselina (DNK) sadrži derivat riboze, deoksiribozu. DNK sadrži nukleotide: adenin, gvanin, citozin i timin. Niz baza određuje genetske informacije. Tri nukleotida u lancu DNK kodiraju jednu aminokiselinu (triplet kod). To. DNK sekcije su geni koji sadrže sve genetske informacije ćelije i služe kao šablon za sintezu ćelijskih proteina.
Glavno svojstvo polinukleotida je sposobnost usmjeravanja reakcija sinteze matriksa (formiranje spojeva - DNK, RNK ili proteina), korištenjem matriksa - specifičnog polinukleotida, a zahvaljujući sposobnosti baza da se međusobno prepoznaju i u interakciji s nekovalentnim veze - ovo je fenomen komplementarnog uparivanja, u kojem se gvanin uparuje sa citozinom, a adenin sa timinom (u DNK) ili uracilom (u RNK).
Komplementarnost je univerzalni princip strukturne i funkcionalne organizacije nukleinskih kiselina i ostvaruje se tokom formiranja DNK i RNK makromolekula tokom replikacije i transkripcije.
Tokom replikacije DNK, novi molekul DNK se gradi na DNK šablonu, tokom transkripcije (formiranje RNK) DNK služi kao šablon, a tokom translacije (sinteza proteina), RNK se koristi kao šablon. U principu, ispostavilo se da je moguć obrnuti proces - konstrukcija DNK na RNK šablonu.
Osim toga, nukleotidi obavljaju još jednu vrlo važnu funkciju u ćeliji: djeluju kao nosioci kemijske energije. Najvažniji (ali ne i jedini) transporter je adenozin trifosfat ili ATP.
U kombinaciji sa drugim hemijskim grupama, nukleotidi su deo enzima. Derivati nukleotida mogu prenijeti određene kemijske grupe s jednog molekula na drugi.
Zagrijavanje, značajna promjena pH vrijednosti, smanjenje jonske snage itd. uzrokuju denaturaciju dvolančane DNK molekule. Termička denaturacija se obično dešava na temperaturi od 80-90C. Moguć je i proces renaturacije molekula DNK (potpuna obnova njegove nativne strukture).
Većina prirodne DNK ima dvolančanu strukturu, linearnu ili kružnu (izuzetak su virusi u kojima se nalazi jednolančana DNK, također linearna ili kružna). U eukariotskoj ćeliji DNK je, osim jezgra, dio mitohondrija i plastida, gdje osigurava autonomnu sintezu proteina. U citoplazmi eukariotskih ćelija pronađeni su analozi DNK bakterijskog plazmida.
Informacije u ćelijama su molekule DNK (kod nekih virusa i bakteriofaga, RNK). Genetske funkcije DNK ustanovljene su 40-ih godina. XX vijek kada proučavamo transformaciju u bakterijama. Ovaj fenomen je prvi opisao 1928. godine F. Griffith dok je proučavao pneumokoknu infekciju kod miševa. Virulentnost pneumokoka određena je prisustvom kapsularnog polisaharida koji se nalazi na površini bakterijskog ćelijskog zida. Virulentne ćelije formiraju glatke kolonije, označene kao S-kolonije (od engleskog smooth - glatko). Avirulentne bakterije, lišene kapsularnog polisaharida kao rezultat mutacije gena, formiraju grube R-kolonije (od engleskog grubo - neravno).
Kao što se vidi iz dijagrama, u jednoj od varijanti eksperimenta Griffith je inficirao miševe mješavinom živih stanica R-soja i mrtvih stanica S-soja. Miševi su uginuli, iako žive bakterije nisu bile infektivne. Žive bakterije izolirane iz mrtvih životinja, kada su posijane na podlogu, formirale su glatke kolonije, budući da su imale polisaharidnu kapsulu. Posljedično, došlo je do transformacije avirulentnih stanica R-soja u virulentne ćelije S-soja. Priroda transformatora je ostala nepoznata.
U 40-im godinama U laboratoriji američkog genetičara O. Averyja prvi put je dobijen DNK preparat pročišćen od proteinskih nečistoća iz ćelija S-soja pneumokoka. Nakon što su ovim lijekom tretirali mutantne stanice R-soja, Avery i njegove kolege (K. McLeod i M. McCarthy) su reproducirali Griffithov rezultat, tj. postignuta transformacija: ćelije su dobile svojstvo virulencije. Tako je utvrđena hemijska priroda supstance koja vrši prenos informacija. Ispostavilo se da je ova supstanca DNK.
Otkriće je bilo prilično neočekivano, jer su do tada naučnici imali tendenciju da pripisuju genetske funkcije proteinima. Jedan od razloga ove greške bio je nedostatak znanja o strukturi molekula DNK. Nukleinske kiseline otkrivene su u jezgru gnojnih ćelija 1869. hemičar I. Mischer, te je proučavan njihov hemijski sastav. Međutim, sve do 40-ih godina. XX vijek naučnici su pogrešno vjerovali da je DNK monoton polimer u kojem se izmjenjuje ista sekvenca od 4 nukleotida (AGCT). Osim toga, nukleinske kiseline su smatrane izuzetno konzervativnim spojevima niske funkcionalne aktivnosti, dok su proteini imali niz svojstava neophodnih za obavljanje genetskih funkcija: polimorfizam, labilnost i prisustvo različitih kemijski aktivnih grupa u njihovim molekulima. I stoga su Avery i njegove kolege počeli da se optužuju za pogrešne zaključke, za nedovoljno pročišćavanje DNK preparata od proteinskih nečistoća. Međutim, poboljšanja u tehnikama pročišćavanja omogućila su potvrdu transformacijske funkcije DNK. Naučnici su uspjeli prenijeti sposobnost formiranja drugih tipova kapsularnih polisaharida u pneumokoke, te također postići transformaciju u druge vrste bakterija za mnoge karakteristike, uključujući otpornost na antibiotike. Značaj otkrića američkih genetičara teško je precijeniti. Služio je kao poticaj za proučavanje nukleinskih kiselina, prvenstveno DNK, u naučnim laboratorijama u mnogim zemljama.
Nakon dokaza o transformaciji u bakterijama, genetske funkcije DNK potvrđene su kod bakteriofaga (bakterijskih virusa). Godine 1952. A. Hershey i S. Chase inficirali su stanice Escherichia coli T2 fagom. Kada se doda u bakterijsku kulturu, ovaj virus se prvo adsorbira na površini ćelije, a zatim ubrizgava svoj sadržaj u nju, što uzrokuje smrt stanice i oslobađanje novih čestica faga. Autori eksperimenta su radioaktivno označili ili DNK T2 faga (32P) ili protein (35S). Čestice faga su pomiješane sa bakterijskim stanicama. Neadsorbirane čestice su uklonjene. Inficirane bakterije su zatim odvojene od praznih ljuski čestica faga centrifugiranjem. Ispostavilo se da je oznaka 35S povezana s ljuskom virusa, koji ostaje na površini ćelije, te stoga virusni proteini ne ulaze u ćeliju. Većina 32P oznake završila je unutar zaražene bakterije. Tako je utvrđeno da su infektivna svojstva bakteriofaga T2 određena njegovom DNK, koja prodire u bakterijsku ćeliju i služi kao osnova za stvaranje novih čestica faga. Ovaj eksperiment je takođe pokazao da fag koristi resurse ćelije domaćina da bi se reprodukovao.
Dakle, početkom 50-ih. XX vijek akumulirano je dovoljno dokaza koji to ukazuju DNK je nosilac genetske informacije. Pored gore navedenih direktnih dokaza, ovaj zaključak je potkrijepljen indirektnim podacima o prirodi lokalizacije DNK u ćeliji, konstantnosti njene količine, metaboličkoj stabilnosti i podložnosti mutagenim efektima. Sve je to potaknulo istraživanje strukture ove molekule.
Pročitajte i druge članke Tema 6 "Molekularne osnove nasljeđa":
Nastavite čitati druge teme u knjizi "Genetika i selekcija. Teorija. Zadaci. Odgovori".







